杨梅花 苗江林 厚富霞 王蕾蕾 程锦涛
(石河子大学农学院,新疆 石河子 832000)
桦木属(Betula)是桦木科(Betulaceae)中分布在新疆的唯一属,我国有31 种及若干变种。新疆是我国西北地区桦木属植被地理分化中心[1],《新疆植物志》[2]中记载有8 个种,其中盐桦(Betula halophila)被列为濒危物种[3]。桦木属植物是组成温带和暖温带森林群落的优势树种[1],具有重要的生态价值和经济价值。
核基因内部转录间隔区(ITS)是植物基因中进化较快的DNA 片段。ITS 序列能提供丰富的信息位点。随着分子技术快速发展,ITS 序列成为重要分子标记,被应用于属及种间分类和系统发育研究[4-6]。国内对于桦木属植物分类和系统发育研究,区域性明显。陈之端[7]从古植物学和地理分布等角度对桦木科植物的起源和演化进行研究;
黄金祥等[8]对河北境内桦木属植物进行了形态分类与地理分布的研究;
刘果厚[9]和侯安燕[10]从植物形态部位对内蒙古和北京地区的桦木属植物分类进行研究;
姜静等[11]利用RAPD 标记技术对东北地区桦树种间亲缘关系进行分析;
梅新娣等[12]应用ITS 序列对盐桦进行了系统发育方面的研究。
目前,从系统分类学对新疆桦木属资源进行研究报道很少。本研究选取新疆桦木属中天山桦(Betula tianschanica)、小叶桦(Betula microphylla)、白 桦(Betula platyphylla)、垂 枝 桦(Betula pendula)、甸生桦(Betula humilis)与盐桦6 种代表植物,以岩桦(Betula calcicola)、高山桦(Betula delavayi)、矮桦(Betula potaninii)和赛黑桦(Betula schmidtii)作为外类群,从分子生物学和形态学角度综合评价新疆桦木属植物的系统分类关系,以期为桦木属植物的系统发育研究提供一定参考依据。
1.1 试验材料
本研究选择新疆桦木属6 个种,其他桦木属10 个种,涵盖《中国植物志》[13]记载的中国境内桦木属2 组及5 亚组植物。16 种桦木属植物ITS序列来自美国国家生物技术信息中心(NCBI)数据库,种名、组/亚组及ITS 序列信息见表1。

表1 桦木属16 种植物ITS 序列信息Table 1 ITS sequences of genus from 16 Betula
1.2 序列分析及系统发育树构建
运用Clustal Omega 软件对桦木属16 种植物ITS 序列进行对比分析,空位被处理为缺失。然后使用MEGA 7.0 软件进行自展分析重复1 000 次,评估简约树中各分支的自展支持率,计算亚组及种间遗传距离,构建最大似然法(ML)系统发育树[14]。
1.3 形态性状的选取及编码
参考《中国植物志》及《新疆植物志》等书籍资料中有关桦木属植物形态性状的记载,选取种间差异大的形态性状,经过SPSS 25.0 软件KMO 检验和Bartlett 球度检验[15],最终选取7 个表型性状作为指标进行聚类分析(表2)。为准确反映出各性状之间的差异程度,采用逻辑顺序赋值量化,确定形态学性状的分析数据。在此基础上,运用SPSS 对统计数据进行主成分及系统聚类分析[16],采用沃德联接法构建形态聚类谱系图。

表2 性状及编码Table 2 Traits and coding
2.1 序列分析及系统发育分析
研究结果显示,桦木属16 种植物的ITS 序列长度范围为604~612 bp,当空位作为缺失处理时,排序后的长度为612 bp。序列位点比对显示共有46 个变异位点,占序列总长的7.52%,其中26 个为简约信息位点。由表3 可知,桦木亚组(Subsect. Betula)与柴桦亚组(Subsect. Fruticosae)、黑桦亚组(Subsect. Dahuricae)之间遗传距离较小,均为0.004 7。硕桦亚组(Subsect. Costatae)内绝对遗传距离较小,为0.003 3。新疆桦木属所在的桦木亚组和柴桦亚组遗传距离较近为0.004 7,桦木亚组内部绝对遗传距离为0.006 0。

表3 桦木属内各亚组间与各亚组内遗传距离Table 3 Genetic distance between and within each subgroup of Betula
由图1 可知,16 种桦木属植物被分为四大支。首先外类群坚桦亚组的岩桦、矮桦、高山桦和赛黑桦聚为F 分支,西桦组的长穗桦(Betula cylindrostachya)、亮叶桦(Betula luminifera)和西桦(Betula alnoides)与硕桦亚组的糙皮桦(Betula utilis)和红桦(Betula albosinensis)各自聚为E 和D 分支。在桦木属ML 系统发育树中,西桦组E 分支支持率达到78%,组内种间序列的自相似度高。新疆桦木属所在的桦木亚组C 分支和柴桦亚组B 分支交错分布,同时该大支中包含有黑桦亚组的黑桦(Betula dahurica)。在桦木亚组分支中,小叶桦与盐桦距离较远,与天山桦距离相对较近。
由图2 可知,新疆桦木属6 种植物中,天山桦与小叶桦、白桦与垂枝桦各自聚集在一小支,遗传距离分别为0.005 0 和0.001 7。甸生桦与盐桦单独聚为一支,遗传距离为0.003 3。

图1 基于ITS 序列分析构建16 种桦木属植物系统发育树Fig. 1 Phylogenetic trees of 16 Betula based on ITS sequence analysis
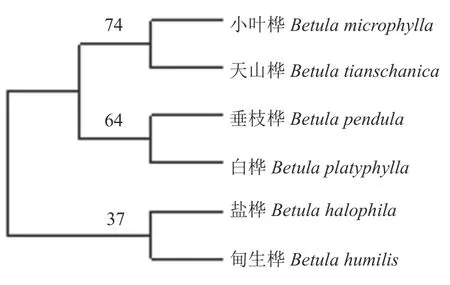
图2 基于ITS 序列分析构建新疆桦木属植物系统发育树Fig. 2 Phylogenetic tree of Betula in Xinjiang based on ITS sequence analysis
2.2 主成分分析及聚类分析
2.2.1 主成分分析
运用SPSS 软件对桦木属16 种植物的7 个形态性状编码进行特征值和方差百分率计算,以特征值大于1 为准则提取主成分。由表4 可知,前3 个主成分累计贡献率达84.5%,第1、2、3 主成分构成比例分别为48.7%、20.5%和15.3%。第1 主成分的特征值为3.407,特征向量中载荷最高且符号为正的性状有生活型和树皮剥裂及纹路,其特征向量为0.816 和0.889。第2 主成分特征值为1.437,从载荷值的大小看,第2 主成分反映叶片形状。第3 主成分特征值为1.071,载荷值最高且符号为正的性状主要是果苞背部被毛,特征向量为0.837。从3 个主成分的特征向量和贡献率分析,生活型、树皮剥裂及纹路是桦木属分类较为重要的性状。

表4 桦木属植物主要性状主成分分析Table 4 Principal component analysis of main characters of Betula
2.2.2 聚类分析
如图3 所示,16 种桦木属植物的形态性状聚类所呈现结果与经典形态分类学拟合度较高。欧氏距离为5 时,长穗桦、亮叶桦和西桦聚为一类,均属西桦组。欧氏距离为4.5 时,外类群坚桦亚组4 个种聚为一支。欧氏距离为3 时,新疆桦木属6 种植物中,桦木亚组的盐桦和柴桦亚组的甸生桦聚为混合类群,而天山桦、小叶桦、垂枝桦和白桦另聚为一个类群。相类似的黑桦亚组的黑桦和硕桦亚组的糙皮桦与红桦聚为一个混合类群。可见,本研究中主成分分析贡献率较高的性状能反映形态分类的主要特点,说明主成分提取方法可行,结果可信。
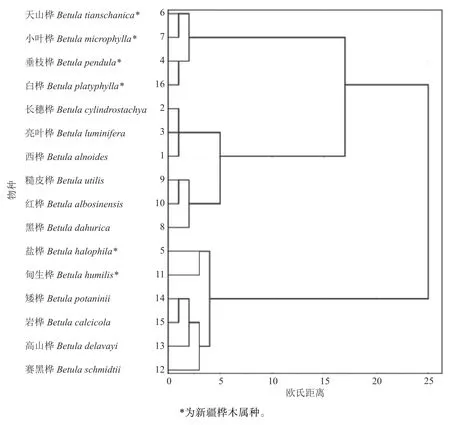
图3 16 种桦木属植物形态性状聚类图Fig. 3 Cluster analysis of 16 Betula
由图4 可知,欧氏距离为5 时,新疆桦木属植物被聚为两大支,分别为单支和混合支。欧氏距离为3 时,柴桦亚组的甸生桦与桦木亚组的盐桦聚为一个混合支,处在树状图的基部位置。另一大支中,桦木亚组的天山桦和小叶桦与垂枝桦和白桦各聚为一单支。总体而言,新疆桦木属的形态性状聚类结果与经典形态分类学中的《新疆植物志》分类结果拟合性较高。

图4 新疆桦木属植物形态性状聚类图Fig. 4 Cluster analysis of Betula in Xinjiang
3.1 新疆桦木属进化关系
在分子系统发育树和形态学聚类结果中,新疆桦木属中天山桦和小叶桦均聚集在一起。地理分布上,小叶桦主要分布在阿尔泰山一带,而天山桦则主要分布在天山山脉[17],但两物种的分布海拔范围重叠程度较高。结合生长环境来看,小叶桦和天山桦都同属于山地森林树种,且从外部形态上均为高大乔木,有着很多共同的特征,如小坚果卵形、树皮灰色,叶菱形等。ITS 分子水平两者的遗传距离为0.005 0,同质性比较高。这一研究结果显示,小叶桦与天山桦在形态学与分子学分类中具有相似性。对于该类桦木属植物,ITS 分子鉴定能够快速准确的进行物种鉴别,这与宫霞等[18]对葡萄科各属的结论和蔡洪月等[19]对杜鹃属的研究结果相一致。
白桦和垂枝桦在2 个ITS 序列系统发育树中均聚为一支,支持率大于60%,并且在桦木属系统发育树中处于较原始的位置,这与姜景民[1]和杨万波等[20]研究桦木属的进化关系结果类似。白桦与垂枝桦在形态学聚类中也聚为一支,与《中国植物志》记载的桦木属分类结果相似,验证了两者在ITS 序列系统发育树中位置的结论。
《中国植物志》分类中甸生桦归属于柴桦亚组,盐桦归属于桦木亚组,而《新疆植物志》中两者被划分在柴桦亚组。本研究中甸生桦和盐桦在形态性状聚类和ITS 序列系统发育树上均聚为一支,说明其形态性状具有一定的共源性,且ITS 序列同源性较高、遗传距离较近。本研究结果支持盐桦与甸生桦分在同一亚组。此外,盐桦与甸生桦都分布在新疆温带草原区,盐桦生活型从小乔木演化为与甸生桦相类似的灌木,反映了相似自然生境对植物的生活型有一定影响[1]。同时,王亚玲等[21]在研究木兰科植物时也有相类似的结果。
3.2 分类群进化及亲缘关系
通过主成分分析可以选择出有效的评价参数和指标,对桦木属形态性状的主成分分析中,生殖器官果苞的形态特征构成第3 主成分,营养器官枝叶性状特征构成第2 主成分,这与从古植物学角度分析桦木属植物系统发育的研究结果一致[10]。
坚桦亚组、硕桦亚组和西桦组在ITS 序列系统发育树中各自聚为一支,且支持率均大于50%。在分子水平上,西桦组与桦木组之间距离较大,绝对遗传距离为0.020 5。桦木组内的坚桦亚组与硕桦亚组之间的遗传距离为0.017 5,而各自亚组内部绝对遗传距离为0.011 3 和0.003 3,证实坚桦亚组与硕桦亚组的亲缘关系较西桦组的亲缘关系近。桦木属内各组、亚组和种间都表现出高的遗传相似性,说明ITS 分子标记鉴定技术的高效性和复杂性[4-5]。演化起源上,西桦组分布于我国西南地区桦木属原始起源中心[1],保留的原始性状比较多,因而与桦木组的坚桦亚组和硕桦亚组遗传特性明显,同时也验证了两者之间遗传距离较远。
桦木亚组、柴桦亚组和黑桦亚组聚为一个混合支,存在不同程度的跨组聚类,与饶龙兵等[22]研究桤木属中出现的情况一致。这可能是因为黑桦亚组和柴桦亚组与桦木亚组的遗传距离均比较小,为0.004 7,表明3 个亚组之间的亲缘关系比较近。研究结果与姜静等[11]发现黑桦亚组和桦木亚组亲缘关系较近的结论一致,但与本研究中形态学聚类结果差异较大。黑桦、糙皮桦和红桦在形态学聚类时聚为一个混合类群,但是它们在经典形态分类学中属于不同亚组种。新疆桦木属植物的研究结果中也出现了类似情况,如甸生桦和盐桦跨亚组聚为一类。推测原因是经典形态分类学主要以形态性状为依据,而植物表型性状受自然环境和人为因素的影响,有时难以阐明种质资源表型性状的差异。特别是桦木属植物为属内多倍体木本植物,不同种之间倍性差异大[5,23],长时间的自然选择和适应环境,使得分子遗传和形态性状上不能很好地吻合,导致属内植物的分类系统十分复杂[24]。ITS 序列在桦木属各亚组间和亚组内遗传距离存在一定间隔,可将各种类进行区分,是桦木属植物较为理想的DNA 条形码候选序列。本研究对新疆桦木属植物的分类及其亲缘关系,从分子系统发育树和形态学聚类进行尝试,今后研究还需结合更多的样本及分子手段进行综合分析。
猜你喜欢桦木亚组遗传非遗传承区域治理(2022年40期)2022-11-27槭叶铁线莲亚组的研究进展园林科技(2021年3期)2022-01-19桦木酸提取、合成及药理作用的研究进展*生物学通报(2020年11期)2020-10-22艾灸神阙穴对不同程度力竭运动大鼠海马区单胺类神经递质的影响❋中国中医基础医学杂志(2020年1期)2020-03-03冠心病患者肠道菌群变化的研究 (正文见第45 页)心电与循环(2020年1期)2020-02-27还有什么会遗传?动漫界·幼教365(小班)(2019年10期)2019-10-28还有什么会遗传动漫界·幼教365(大班)(2019年10期)2019-10-28还有什么会遗传?动漫界·幼教365(中班)(2019年10期)2019-10-28黑漆桦木椅三联生活周刊(2017年9期)2017-03-03黄龙山白桦桦木醇与桦木酸含量研究西北植物学报(2015年9期)2015-07-05